
California and Western Medicine
Articles
CALIFORNIA
AND
WESTERN MEDICINE
VOLUME XXXV - DECEMBER, 1931 - NO. 6
OBSERVATIONS ON BACILLUS TYPHOSUS IN ITS FILTERABLE STATE
A PRELIMINARY COMMUNICATION
By ARTHUR ISAAC KENDALL, PH.D. Chicago, Illinois
and
ROYAL RAYMOND RIFE, PH.D.
San Diego, California
From:
The Rife Research Laboratory, San Diego, California; The Laboratory or Research Bacteriology, Northwestern University Medical School, Chicago, Illinois; and The Pathological Laboratory of the Pasadena Hospital, Pasadena, California.
Presented at a meeting of the Bacteriological Section or the Los Angeles Clinical and Pathological Society, November 20, 1931.
It seems improbable that viable bacteria in the filterable state have ever been unequivocally seen. Nevertheless, the theoretical and practical importance of filterable forms of bacteria in theoretical and applied biology cannot be denied.
The organism selected for these experiments was the well known Rawlings strain of B. typhosus. The immediate history of the culture used is as follows:
October 29, 1931. An agar slant was made of a thrice-plated culture of B. typhosus, Rawlings strain. (Editor's Note: This agar slant was made in the Laboratory of Research Bacteriology, Northwestern University Medical School, Chicago, Illinois.)
November 2, 4 p. m. Inoculated six cubic centimeters of K (protein) Medium 2 from the agar slant culture.
November 3, 10 a. m. Filtered this culture in K Medium of November 2, through a Berkefeld "N" filter. (The culture was diluted with four volumes of sterile physiological saline solution; the vacuum used was less than four inches of water; the total time of filtration was less than ten minutes.)
November 3. One drop of filtrate, representing one-fifth drop of the original culture, was introduced into six cubic centimeters of K Medium. Incubated at 37 degrees centigrade. The filtrate was also tested for purity as follows: (1) cultural reactions; (2) sugar fermentation reactions; (3) agglutination with specific typhoid serum. All were typical.
November 5. The forty-eight-hour culture of November 3 in K Medium was filtered, as above, through a Berkefeld "N" filter. One drop of the filtrate was added to six cubic centimeters of K Medium and incubated at 37 degrees centigrade.
November 6. The twenty-four-hour culture of November 5, which grew well, was again filtered. This time the finest Berkefeld filter, "W," was used. As before, a drop of this filtrate was added to six cubic centimeters of K medium and incubated at 37 degrees centigrade. Growth was abundant November 7.
November 9. The culture was again transferred to K Medium.
November 12. Still another culture was made, in every instance using three loops of culture for the inoculum.
It is worthy of note that this thrice filtered culture of B. typhosus grew quite readily in K Medium as above outlined: after the second filtration it failed to grow in peptone broth. In other words, the organism having become filterable and accustomed to protein media (proteophilic) lost its ability to grow in ordinary peptone containing nutrient broth.
The cultures of November 9 and November 12 were examined under the microscope and there were no discernible bacilli, although the cultures were markedly turbid. Darkfield illumination revealed very small, actively motile granules, and direct observation of these with the oil emersion lens confirmed the presence of these motile granules, without, however, affording any indication of their structure; Therefore, these granules for obvious reasons could not be unequivocally diagnosed as the filterable form of the bacillus.
The organism selected for these experiments was the well known Rawlings strain of B. typhosus. The immediate history of the culture used is as follows:
November 6. The twenty-four-hour culture of November 5, which grew well, was again filtered. This time the finest Berkefeld filter, "W," was used. As before, a drop of this filtrate was added to six cubic centimeters of K medium and incubated at 37 degrees centigrade. Growth was abundant November 7.
In this viable, filtered state the culture was taken to Pasadena, California, and, through the instrumentality of Dr. Milbank Johnson, the cooperation of Dr. Alvin G. Foord, and the courtesy of the Pasadena Hospital, the necessary space and equipment for mounting the microscope and continuing the cultures were made available. The subsequent developments, which are the immediate subject of this discussion, are as follows:
When a culture of B. typhosus in the filterable state, grown as above indicated in K Medium, was examined with this micropolarimeter, it was observed that the plane of polarization of the light passing through the culture was deviated plus 4.8 degrees, with the simultaneous appearance of a definite blue spectrum. With this observation in mind, the culture was next studied with the Rife microscope at 5000 diameters.
The double wedge quartz prism referred to above was set by means of the vernier to minus 4.8 degrees (The reason for setting the quartz wedge in the reverse direction will be discussed in another place). Examined in this polarized light, this thrice filtered culture of B. typhosus cultivated in K (protein) Medium showed small, oval granules, many of them quite actively motile. These motile granules when in true focus appeared as bright turquoise-blue bodies, which contrast strikingly, both in color and in their active motion, with the noncolored, nonmotile débris of the medium.
These observations were repeated eight times, using in each instance growth of the filterable organisms in K Medium. The cultures examined were both twenty-four and forty-eight hours old. The qualitative results were always the same, namely, the occurrence of small, oval, actively motile, turquoise-blue bodies in the cultures and the absence of these small, oval, actively motile, turquoise-blue bodies in the uninoculated control K Media.
There is another even more direct procedure for establishing the identity of these small, oval, motile, turquoise-blue bodies. It has been shown in previous communications 3 that agar cultures, or better, broth cultures of B. typhosus inoculated into K Medium, become filterable within eighteen hours' growth at 37 degrees centigrade. It should follow, inasmuch as not all of the bacilli appear to become filterable under these conditions, that at least some of the bacilli should have similar turquoise-blue granules within their substance if they are indeed passing to the filterable state. Also the free swimming filterable forms, the small, oval, motile, turquoise-blue bodies described above, should be simultaneously present.
Dark field examination of such a culture eighteen hours old revealed unchanged, actively motile bacilli, bacilli with granules within their substance, and free swimming, actively motile granules. This culture examined in the Rife microscope with the quartz prism set at minus 4.8 degrees and with 5000 diameters magnification, showed very clearly the three types of organisms just described, namely:
First, unchanged bacilli: These were relatively long, actively motile, and almost devoid of color.
November 16. The cultures of November 12, made in Chicago, were transferred to fresh K Medium and incubated at 37 degrees centigrade overnight.
November 17. The Rife microscope was installed and the first cultures, those inoculated November 16, were examined. The preliminary observations of these cultures were made with a polarizing microscope with a spectroscopic attachment. It should be borne in mind that the entire optical system of this micropolarimeter was of quartz. A one-eighteenth-inch apochromatic oil immersion lens was used, with a 20x quartz ocular.
When a culture of B. typhosus in the filterable state, grown as above indicated in K Medium, was examined with this micropolarimeter, it was observed that the plane of polarization of the light passing through the culture was deviated plus 4.8 degrees, with the simultaneous appearance of a definite blue spectrum. With this observation in mind, the culture was next studied with the Rife microscope at 5000 diameters.
The double wedge quartz prism referred to above was set by means of the vernier to minus 4.8 degrees (The reason for setting the quartz wedge in the reverse direction will be discussed in another place). Examined in this polarized light, this thrice filtered culture of B. typhosus cultivated in K (protein) Medium showed small, oval granules, many of them quite actively motile. These motile granules when in true focus appeared as bright turquoise-blue bodies, which contrast strikingly, both in color and in their active motion, with the noncolored, nonmotile débris of the medium.
These observations were repeated eight times, using in each instance growth of the filterable organisms in K Medium. The cultures examined were both twenty-four and forty-eight hours old. The qualitative results were always the same, namely, the occurrence of small, oval, actively motile, turquoise-blue bodies in the cultures and the absence of these small, oval, actively motile, turquoise-blue bodies in the uninoculated control K Media.
There is another even more direct procedure for establishing the identity of these small, oval, motile, turquoise-blue bodies. It has been shown in previous communications 3 that agar cultures, or better, broth cultures of B. typhosus inoculated into K Medium, become filterable within eighteen hours' growth at 37 degrees centigrade. It should follow, inasmuch as not all of the bacilli appear to become filterable under these conditions, that at least some of the bacilli should have similar turquoise-blue granules within their substance if they are indeed passing to the filterable state. Also the free swimming filterable forms, the small, oval, motile, turquoise-blue bodies described above, should be simultaneously present.
Dark field examination of such a culture eighteen hours old revealed unchanged, actively motile bacilli, bacilli with granules within their substance, and free swimming, actively motile granules. This culture examined in the Rife microscope with the quartz prism set at minus 4.8 degrees and with 5000 diameters magnification, showed very clearly the three types of organisms just described, namely:
First, unchanged bacilli: These were relatively long, actively motile, and almost devoid of color.
Second, long, actively motile bacilli, each with a rather prominent granule at one end. The granule in such an organism was turquoise blue, reminiscent in size, shape, and color of the small, oval, actively motile, turquoise-blue granules found in the protein medium (K Medium) where, it will be recalled, no formed (rod shaped) bacteria could be demonstrated. These bacilli having the turquoise-blue granules were colored only at the granule end, the remainder of the rod being nearly colorless, in this respect corresponding to the unchanged (nonfilterable) bacilli just mentioned.
Third, free swimming, small, oval, actively motile, turquoise-blue granules, precisely similar, apparently, in size, shape, and color to those seen in the granulated bacilli just described.
Laboratory of Medical Research, Northwestern University Medical School, 303 Chicago Avenue, Chicago, Illinois.
Rife Research Laboratory, 712 Electric Building, San Diego.
Laboratory of Medical Research, Northwestern University Medical School, 303 Chicago Avenue, Chicago, Illinois.
Rife Research Laboratory, 712 Electric Building, San Diego.
From the two facts thus far arrived at, namely, that the small, oval, turquoise-blue bodies were actively motile and also that they were cultivable from K Medium to K Medium, it is surmised that these small, oval, motile turquoise-blue bodies are indeed the filterable forms of the B. typhosus.
From the fact that these small, oval, turquoise-blue bodies could be seen both in the parent rod and free swimming in the medium, it is assumed that these small, oval, actively motile, turquoise-blue bodies are indeed the filterable form of B. typhosus.
Laboratory of Medical Research, Northwestern University Medical School, 303 Chicago Avenue, Chicago, Illinois.
Rife Research Laboratory, 712 Electric Building, San Diego.
REFERENCES
1. James ,4. Patten Lecture, Northwestern University Bulletin, Vol. 32, No. 5 (September 28), 1931.
2. Northwestern University Medical School Bulletin, Vol. 32, No. 8, (October 19), 1931, for full details.
3. Op. cit.
2. Northwestern University Medical School Bulletin, Vol. 32, No. 8, (October 19), 1931, for full details.
3. Op. cit.
Es scheint unwahrscheinlich, dass lebensfähige Bakterien im filtrierbaren Zustand jemals eindeutig gesehen wurden. Dennoch kann die theoretische und praktische Bedeutung der filtrierbaren Formen von Bakterien in der theoretischen und angewandten Biologie nicht geleugnet werden.
Durch die gleichzeitige Verfügbarkeit des Rife-Mikroskops, einem Instrument, das sehr hohe Vergrößerung mit koordinierter Auflösungskraft kombiniert, und einem einfachen Verfahren zur Induktion des filtrierbaren Zustandes in Bakterien nach Belieben1 , bot sich in jüngster Zeit die Möglichkeit, Organismen in diesem bisher illusorischen Zustand tatsächlich nachzuweisen.
Zwei Merkmale des Rife-Mikroskops, die an anderer Stelle im Detail vorgestellt werden, müssen hier ausdrücklich erwähnt werden. Zum einen ist das gesamte optische System, also nicht nur die Linsen, sondern auch die Beleuchtungseinheit, aus Quarzglas gefertigt. Zusätzlich ist ein Doppelkeil-Quarzprisma zwischen der Beleuchtungseinheit und dem Quarz-Abbé-Kondensor montiert. Letzterer ist um 360 Grad mit einem Nonius drehbar und ermöglicht so ein leicht steuerbares polarisiertes Licht in jedem gewünschten Winkel. Der Import dieser Polarisationseinheit wird später besprochen. Da dieses Mikroskop von 5.000 auf 17.000 Durchmesser vergrößert, ist es natürlich sehr notwendig, es auf einem unbeweglichen Fundament zu montieren.
Der für diese Experimente ausgewählte Organismus war der bekannte Rawlings-Stamm von B. typhosus. Die unmittelbare Geschichte der verwendeten Kultur ist wie folgt:
29. Oktober 1931. Eine Agarschräge wurde aus einer dreifach plattierten Kultur von B. typhosus, Rawlings-Stamm, hergestellt. (Anmerkung des Herausgebers: Dieser Agar wurde im Laboratorium für Forschungsbakteriologie der Northwestern University Medical School, Chicago, Illinois, hergestellt).
2. November, 16.00 Uhr. Es wurden sechs Kubikzentimeter K (Protein) Medium 2 aus der Agar Slant Kultur geimpft.
3. November, 10 Uhr morgens. Filtrierte diese Kultur in K (Protein) Medium 2 vom 2. November durch einen Berkefeld "N"-Filter. (Die Kultur wurde mit vier Volumina steriler physiologischer Kochsalzlösung verdünnt; das verwendete Vakuum betrug weniger als vier Zentimeter Wasser; die Gesamtzeit der Filtration betrug weniger als zehn Minuten).
3. November. Ein Tropfen Filtrat, der ein Fünftel des Tropfens der ursprünglichen Kultur ausmacht, wurde in sechs Kubikzentimeter K-Medium gegeben. Die Inkubation erfolgte bei 37 Grad Celsius. Das Filtrat wurde auch wie folgt auf seine Reinheit getestet: (1) Kulturreaktionen; (2) Zuckerfermentationsreaktionen; (3) Agglutination mit spezifischem Typhus-Serum. Alle waren typisch.
5. November. Die achtundvierzigstündige Kultur vom 3. November in K-Medium wurde wie oben durch einen Berkefeld-"N"-Filter gefiltert. Ein Tropfen des Filtrats wurde sechs Kubikzentimetern K-Medium hinzugefügt und bei 37 Grad Celsius inkubiert.
6. November. Die vierundzwanzigstündige Kultur vom 5. November, die gut gewachsen ist, wurde erneut gefiltert. Diesmal wurde der feinste Berkefeld-Filter "W" verwendet. Wie zuvor wurde ein Tropfen dieses Filtrats auf sechs Kubikzentimeter K-Medium gegeben und bei 37 Grad Celsius bebrütet. Das Wachstum war am 7. November reichlich vorhanden.
9. November. Die Kultur wurde erneut in K-Medium überführt.
12. November. Eine weitere Kultur wurde angelegt, wobei jeweils drei Kulturschleifen für das Inokulum verwendet wurden.
Es ist bemerkenswert, dass diese dreimal gefilterte Kultur von B. typhosus, wie oben beschrieben, recht leicht in K Medium wuchs: Nach der zweiten Filtration wuchs sie nicht mehr in Peptonbouillon. Mit anderen Worten, nachdem der Organismus filtrierbar geworden war und sich an Proteinmedien (proteophil) gewöhnt hatte, verlor er seine Fähigkeit, in gewöhnlicher peptonhaltiger Nährbouillon zu wachsen.
Die Kulturen vom 9. und 12. November wurden unter dem Mikroskop untersucht, und es waren keine Bazillen zu erkennen, obwohl die Kulturen deutlich getrübt waren. Die Dunkelfeldbeleuchtung zeigte sehr kleine, aktiv bewegliche Körnchen, deren direkte Beobachtung mit der Ölaustrittslinse das Vorhandensein dieser beweglichen Körnchen bestätigte, ohne jedoch einen Hinweis auf ihre Struktur zu geben; daher konnten diese Körnchen aus offensichtlichen Gründen nicht eindeutig als die filtrierbare Form des Bazillus diagnostiziert werden.
Der für diese Experimente ausgewählte Organismus war der bekannte Rawlings-Stamm von B. typhosus. Die unmittelbare Geschichte der verwendeten Kultur ist wie folgt:
6. November. Die vierundzwanzigstündige Kultur vom 5. November, die gut gewachsen ist, wurde erneut gefiltert. Diesmal wurde der feinste Berkefeld-Filter "W" verwendet. Wie zuvor wurde ein Tropfen dieses Filtrats auf sechs Kubikzentimeter K-Medium gegeben und bei 37 Grad Celsius bebrütet. Das Wachstum war am 7. November reichlich vorhanden.
In diesem lebensfähigen, gefilterten Zustand wurde die Kultur nach Pasadena, Kalifornien, gebracht, und mit Hilfe der Instrumente von Dr. Milbank Johnson, der Mitarbeit von Dr. Alvin G. Foord und der Höflichkeit des Krankenhauses von Pasadena wurden der erforderliche Raum und die Ausrüstung für die Anbringung des Mikroskops und die Fortsetzung der Kulturen zur Verfügung gestellt. Die nachfolgenden Entwicklungen, die unmittelbar Gegenstand dieser Diskussion sind, sind wie folgt:
16. November. Die Kulturen vom 12. November, die in Chicago hergestellt wurden, wurden in frisches K-Medium überführt und über Nacht bei 37 Grad Celsius bebrütet.
17. November. Das Rife-Mikroskop wurde installiert und die ersten Kulturen, die am 16. November geimpft wurden, wurden untersucht. Die vorläufigen Beobachtungen dieser Kulturen wurden mit einem Polarisationsmikroskop mit spektroskopischem Aufsatz durchgeführt. Man sollte bedenken, dass das gesamte optische System dieses Mikropolarimeters aus Quarz bestand. Es wurde eine apochromatische Ölimmersionslinse von einem achtzehnten Zoll mit einem 20fachen Quarzokular verwendet.
Bei der Untersuchung einer Kultur von B. typhosus im filtrierbaren Zustand, die wie oben in K-Medium gezüchtet wurde, wurde mit diesem Mikropolarimeter beobachtet, dass die Polarisationsebene des durch die Kultur hindurchtretenden Lichts um 4,8 Grad plus 4,8 Grad abgelenkt wurde, bei gleichzeitigem Auftreten eines bestimmten blauen Spektrums. Mit dieser Beobachtung im Hinterkopf wurde die Kultur als nächstes mit dem Rife Mikroskop bei 5000 Durchmessern untersucht.
Das oben erwähnte Doppelkeil-Quarzprisma wurde mit Hilfe des Nonius auf minus 4,8 Grad eingestellt (Der Grund für die Einstellung des Quarzkeils in umgekehrter Richtung wird an anderer Stelle diskutiert). Diese dreimal gefilterte Kultur von B. typhosus, die in K (Protein) Medium kultiviert wurde, zeigte bei der Untersuchung in diesem polarisierten Licht kleine, ovale Körnchen, von denen viele recht aktiv beweglich waren. Diese beweglichen Körnchen erschienen im echten Fokus als helle türkisblaue Körper, die sowohl in der Farbe als auch in ihrer aktiven Bewegung in auffälligem Kontrast zu den nicht gefärbten, nicht beweglichen Débris des Mediums standen.
Diese Beobachtungen wurden acht Mal wiederholt, wobei jeweils das Wachstum der filtrierbaren Organismen im K-Medium verwendet wurde. Die untersuchten Kulturen waren sowohl vierundzwanzig als auch achtundvierzig Stunden alt. Die qualitativen Ergebnisse waren immer die gleichen, nämlich das Auftreten kleiner, ovaler, aktiv beweglicher, türkisblauer Körper in den Kulturen und das Fehlen dieser kleinen, ovalen, aktiv beweglichen, türkisblauen Körper in den nicht geimpften Kontroll-K-Medien.
Es gibt ein weiteres, noch direkteres Verfahren zur Feststellung der Identität dieser kleinen, ovalen, beweglichen, türkisblauen Körper. In früheren Mitteilungen 3 wurde gezeigt, dass Agar-Kulturen, oder besser gesagt, Brühenkulturen von B. typhosus, die in K-Medium geimpft werden, innerhalb von achtzehn Stunden Wachstum bei 37 Grad Celsius filtrierbar werden. Da nicht alle Bazillen unter diesen Bedingungen filtrierbar zu sein scheinen, sollten zumindest einige der Bazillen ähnliche türkisblaue Körnchen in ihrer Substanz haben, wenn sie tatsächlich in den filtrierbaren Zustand übergehen. Auch die frei schwimmenden filtrierbaren Formen, die kleinen, ovalen, beweglichen, türkisblauen Körper, die oben beschrieben wurden, sollten gleichzeitig vorhanden sein.
Die Dunkelfelduntersuchung einer solchen achtzehn Stunden alten Kultur ergab unveränderte, aktiv bewegliche Bazillen, Bazillen mit Körnern in ihrer Substanz und frei schwimmende, aktiv bewegliche Körner. Diese im Rife-Mikroskop untersuchte Kultur mit dem auf minus 4,8 Grad eingestellten Quarzprisma und mit einer Vergrößerung von 5000 Durchmessern zeigte sehr deutlich die drei soeben beschriebenen Organismentypen, nämlich
Erstens, unveränderte Bazillen: Diese waren relativ lang, aktiv beweglich und fast farblos.
Zweitens, lange, aktiv bewegliche Bazillen, die jeweils an einem Ende ein ziemlich hervorstehendes Granulat aufweisen. Die Körnchen in einem solchen Organismus waren türkisblau und erinnerten in Größe, Form und Farbe an die kleinen, ovalen, aktiv beweglichen, türkisblauen Körnchen im Proteinmedium (K-Medium), wo, wie wir uns erinnern werden, keine gebildeten (stäbchenförmigen) Bakterien nachgewiesen werden konnten. Diese Bazillen mit den türkisblauen Körnchen waren nur am Ende der Körnchen gefärbt, der Rest des Stäbchens war nahezu farblos, was in dieser Hinsicht den eben erwähnten unveränderten (nicht filtrierbaren) Bazillen entspricht.
Drittens, frei schwimmende, kleine, ovale, aktiv bewegliche, türkisblaue Körnchen, die in Größe, Form und Farbe genau denen der eben beschriebenen granulierten Bazillen entsprechen.
Laboratorium für medizinische Forschung, Northwestern University Medical School, 303 Chicago Avenue, Chicago, Illinois.
Rife Research Laboratory, 712 Electric Building, San Diego.
Aus den beiden bisher ermittelten Tatsachen, nämlich dass die kleinen, ovalen, türkisblauen Körper aktiv beweglich waren und dass sie von K Medium bis K Medium kultivierbar waren, wird vermutet, dass diese kleinen, ovalen, beweglichen türkisblauen Körper tatsächlich die filtrierbaren Formen des B. typhosus sind.
Aus der Tatsache, dass diese kleinen, ovalen, türkisblauen Körper sowohl in der Elternrute als auch frei schwimmend im Medium zu sehen waren, wird angenommen, dass diese kleinen, ovalen, aktiv beweglichen, türkisblauen Körper tatsächlich die filtrierbare Form des B. typhosus sind.
Laboratorium für medizinische Forschung, Northwestern University Medical School, 303 Chicago Avenue, Chicago, Illinois.
Rife Research Laboratory, 712 Electric Building, San Diego.
REFERENZEN
1. James ,4. Patten Lecture, Northwestern University Bulletin, Vol. 32, No. 5 (September 28), 1931.
2. Northwestern University Medical School Bulletin, Vol. 32, No. 8, (October 19), 1931, for full details.
3. Op. cit.
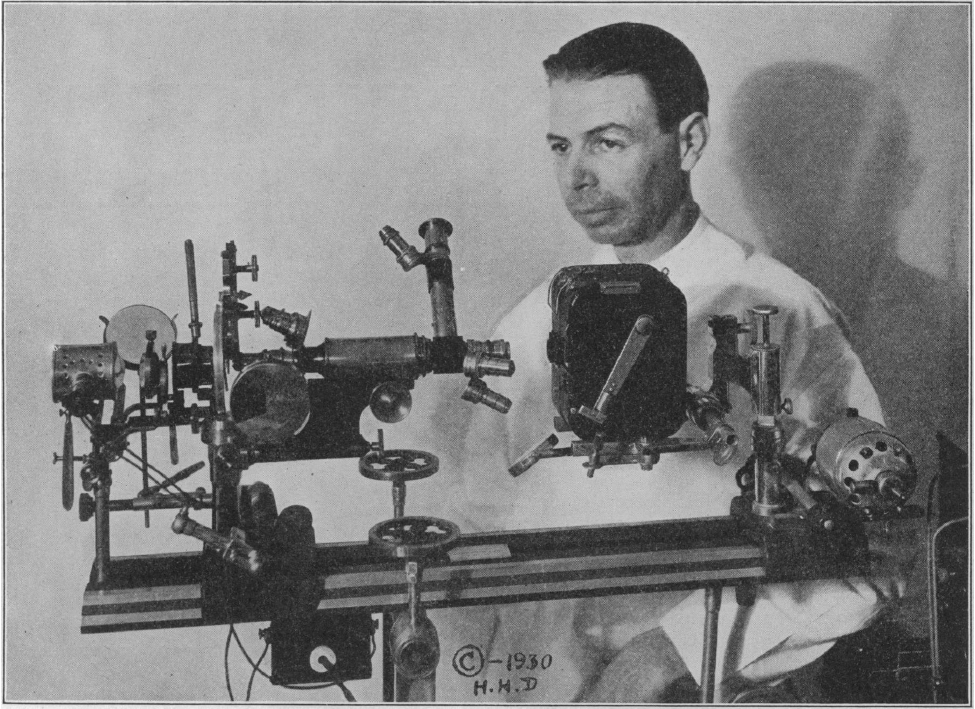
Fig. 1.--Photograph of the microscope and its inventor. Royal Raymond Rife. Ph. D. In the illustration the source of illumination is to the extreme left, the light passing through the substation condenser and then through the optical system. The vertical tube is the observation tube. The three lenses are directed toward the camera. The camera is a special stop motion camera for standard films. Beyond the camera is the motor to drive it. The stage on which the instrument is placed is so arranged that the microscope can be tilted through any axis from horizontal to vertical.
Abb. 1: Foto des Mikroskops und seines Erfinders. Königlicher Raymond Rife. Ph. D. In der Abbildung ist die Beleuchtungsquelle ganz links, das Licht geht durch den Kondensor der Unterstation und dann durch das optische System. Der vertikale Tubus ist der Beobachtungstubus. Die drei Linsen sind auf die Kamera gerichtet. Die Kamera ist eine spezielle Stop-Motion-Kamera für Standardfilme. Hinter der Kamera befindet sich der Motor, der die Kamera antreibt. Der Tisch, auf dem das Instrument steht, ist so angeordnet, dass das Mikroskop um jede Achse von der Horizontalen in die Vertikale gekippt werden kann.
Abb. 1: Foto des Mikroskops und seines Erfinders. Königlicher Raymond Rife. Ph. D. In der Abbildung ist die Beleuchtungsquelle ganz links, das Licht geht durch den Kondensor der Unterstation und dann durch das optische System. Der vertikale Tubus ist der Beobachtungstubus. Die drei Linsen sind auf die Kamera gerichtet. Die Kamera ist eine spezielle Stop-Motion-Kamera für Standardfilme. Hinter der Kamera befindet sich der Motor, der die Kamera antreibt. Der Tisch, auf dem das Instrument steht, ist so angeordnet, dass das Mikroskop um jede Achse von der Horizontalen in die Vertikale gekippt werden kann.

Fig. 2.--Arthur Isaac Kendall, Ph. D., Director of Medical Research. Northwestern University Medical co-author with Royal Raymond Rife, Ph. D., of the paper on "Observations on Bacillus Typhosus in Its Filterable State."
Abb. 2: Dr. Arthur Isaac Kendall, Direktor der medizinischen Forschung. Northwestern University Medical Co-Autor mit Royal Raymond Rife, Ph. D., der Arbeit "Beobachtungen zum Bacillus Typhosus in seinem filtrierbaren Zustand".
Abb. 2: Dr. Arthur Isaac Kendall, Direktor der medizinischen Forschung. Northwestern University Medical Co-Autor mit Royal Raymond Rife, Ph. D., der Arbeit "Beobachtungen zum Bacillus Typhosus in seinem filtrierbaren Zustand".